Bitkilerde Taşıma
En küçük otsu bitkiden en yüksek ağaçlara kadar her bitki, fotosentez için gerekli maddeleri hücrelerine sağlamak ve fotosentez sonucu meydana gelen ürünleri gerekli bölgelere taşımak zorundadır. Basit yapılı bitkilerde bu iş ozmos, difüzyon ve aktif taşıma ile gerçekleştirildiği için taşıma sistemi bulunmaz. Ancak bitkilerin çoğunda su ve minerallerin hücreden hücreye geçerek kökten yapraklara kadar pasif ve aktif taşıma ile ulaştırılması, yine yapraklarda oluşan organik bileşiklerin benzer yolla gerekli yerlere iletilmesi imkânsızdır. Bu tür bitkilerde problem taşıma sistemi (iletim dokuları) ile çözülmüştür. Daha önce de anlatıldığı gibi su ve çözünmüş minerallerin kökten yapraklara kadar taşınmasını ksilem adı verilen iletim dokusu, yapraklardaki fotosentez ürünlerinin gerekli yerlere taşınmasını da floem adı verilen iletim dokusu sağlar.
Bitkilerde Su ve Minerallerin Taşınması
Bitkilerde suyun çok büyük bir bölümü kök uçlarından alınır. Yapılan araştırmalar en fazla ve en hızlı su alımının kök ucunun emici tüy bölgesinde olduğunu göstermiştir. Emici tüyler epidermis hücrelerinin toprak içine doğru uzantıları olup toprak parçacıklarıyla temas yüzeyini artırırlar. Bu yapılar çok ince oldukları için, topraktaki en küçük delikçiklere bile girerek su ve minerallerin alınmasını sağlayabilirler.
Emici tüyler ortalama 4-5 gün yaşar ve sürekli olarak yeni tüyler oluşturulur. Bir çavdar bitkisinde emici tüylerin sayısı 14 milyara kadar çıkabilir ve bunların oluşturduğu toplam yüzey alanı 400 m2 den daha fazla bir yer kaplar. Emici tüyden içeri giren su ve mineraller korteksi geçerek endodermise gelir. Daha sonra merkezi silindirdeki ksilemin içine girer.
Bitki kök hücrelerinin zarlarında bulunan taşıyıcı proteinler, mineral besin elementlerinin alınmasında çok önemli rol oynar. Eğer ihtiyaç duyulan iyonun topraktaki derişimi, hücredekinden fazla ise ve hücre zarında bulunan proteinlerin oluşturduğu uygun kanallar varsa, iyonlar ATP harcanmadan içeri girer. Yani iyonların hücre içine girişi kolaylaştırılmış difüzyonla olur. Ancak ihtiyaç duyulan mineral iyonlarının topraktaki derişimi hücredekinden genellikle daha azdır. Bu durumda mineraller ATP kullanılarak, yani aktif taşımayla hücre içine alınır.
Minerallerin kökteki ksileme ulaşması için iki yol vardır. Birinci yolda su ve mineraller hücreye girmeden, hücreler arası boşluklardan geçerek; ikinci yolda ise hücreden hücreye uzanan sitoplâzma bağlantıları aracılığıyla taşınır.
Suyun Gövdede Taşınmasını Sağlayan Mekanizmalar
Su ve suda çözünmüş minerallerin kökten yapraklara kadar taşınmasını ksilem adı verilen iletim dokusu sağlar, «şilemde taşıma aşağıdan yukarıya doğru tek yönlüdür.
Bitkilerde suyun ksilemde taşınmasını açıklamak amacıyla araştırmacılar tarafından çeşitli mekanizmalar ileri sürülmüştür. Ksilem özsuyu, bazı ağaçlarda 100 m’den daha fazla yüksekliğe ulaşabilmektedir. Suyun hareket edebilmesini temel olarak iki mekanizma sağlayabilir. Bunlardan birincisi suyun köklerden yukarı doğru itilmesidir ve bu olay “kök basıncı” olarak adlandırılır. İkincisi de suyun yapraklardan yukarı doğru çekilmesidir. Bu mekanizma ise “terleme - çekim teorisi” ya da “kohezyon - gerilim teorisi” adını alır. Uzun bir süre suyun hareketinde odun borularının “kılcallık” özelliği de önemli bir faktör olarak düşünülmüş, ancak daha sonra bunun çok yetersiz kaldığı anlaşılmıştır.
Kök Basıncı: Bitki kökleri tarafından toprak çözeltisinden alınan mineraller merkezi silindire taşınır. Merkezi silindiri çeviren endodermis tabakası bu iyonların geri çıkmalarını engeller. Merkezi silindirde biriken çözünmüş mineraller, bu bölgenin su potansiyelini düşürür. Bu durumda ozmotik kurallara göre dışarıdaki su kökün içine girer ve ksileme doğru ilerler. Bitki köküne hızla giren suyun oluşturduğu hidrostatik basınç kök basıncını meydana getirir. Kök basıncı, suyun ksilemde hareketini sağlar.
Kök basıncı ile suyun hareketi basit bir deneyle gösterilebilir. Saksıda yetiştirilen domates bitkisi iyice sulandıktan sonra gövdesinden kesilir ve kesik yüzeye kauçuk boru yardımıyla içerisinde biraz su bulunan ince bir cam boru takılırsa, kısa bir süre geçince cam boruda suyun yukarı doğru yükseldiği görülür.
Birçok bitkide kök basıncı, gutasyon (damlama) adı verilen bir olaya yol açar. Bitkilerin yapraklarından sıvı şekilde su kaybetmeleri gutasyon olarak adlandırılmaktadır. Yapraklarda damlama, "hidatod'’ adı verilen deliklerden gerçekleşir. Yaprak kenarında ve ucunda damlama sonucu ortaya çıkan suyu “çiğ” ile karıştırmamak gerekir. Çiğ, yaprağın kenar ve ucunda değil üst yüzeyinde oluşur. Damlama olayı çoğunlukla su alımının hızlı ve terlemenin az olduğu koşullarda belirginleşir.
Toprakta yeterince su bulunduğu ve geceleri nem düzeyi yüksek olduğu zaman ksilemdeki su, kök basıncının etkisiyle hidatoddan damlalar halinde dışarı çıkar. Çıkan bu su, saf su olmayıp çözünmüş maddeler içermektedir.
Kök basıncı suyun sadece birkaç metre yükselmesini sağladığı için dev sekoyalar gibi uzun boylu ağaçlarda, suyu yukarıya doğru itmeye yetmez. Ayrıca kök basıncının hiç olmadığı durumlarda bile bitkilerde su taşınabilmektedir. Bu durum su taşınmasının sadece kök basıncı ile açıklanamayacağını ortaya koymaktadır.
Terleme - Çekim Teorisi: Çoğunlukla hidrojen bağları sayesinde moleküllerin bir arada tutulması kohezyon olarak adlandırılır. Farklı moleküller arasındaki çekim kuvveti de adezyon adını alır. Terleme çekim teorisine göre; su moleküllerinin birbirine bağlanma (kohezyon) gücünün, içinde taşındıkları iletim borularına bağlanma (adezyon) gücünün ve terleme sonucu bitkinin yukarı kısımlarında oluşan emme gücünün yardımlarıyla ksilem içerisinde su yukarı doğru taşınmaktadır.
Yaprağın mezofil hücrelerinde fotosentez ve terlemeyle su kaybedilmesi sonucu ozmotik basınç artışı olur. Bunun sonucu bitkinin üst kısımlarında bir emme (çekme) kuvveti ortaya çıkar. Su molekülleri hidrojen bağları ile birbirine bağlı olduğundan, kopmayan bir sütun halinde ksilemden yukarı doğru çekilir. Bu arada kaybedilen suyun yerine de kök emici tüyleriyle topraktan yeniden su emilir.
Terleme - çekim teorisi basit bir deneyle gösterilebilir. İçinde renkli su olan bir behere, içi suyla dolu ince uzun bir cam boru daldırılır. Cam borunun ucuna ıslak bir sünger konulur. Cam borudaki su sütunu; alttan beherdeki renkli suyla, üstten de ıslak süngerle temas halindedir. Süngerdeki su buharlaştıkça su sütunu yukarı doğru çekildiği için cam borudaki renkli sıvının yükseldiği görülür.
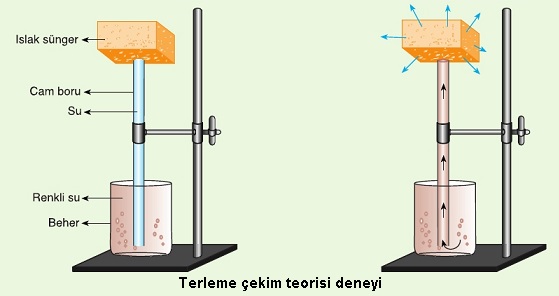
Yukarıdaki deneyde, beherdeki suyu toprak suyuna, cam boruyu ksileme ve ıslak süngeri de buharlaşmanın çok olduğu yaprak mezofil hücrelerinin yüzeylerine benzetebiliriz. Topraktaki su ile yapraktaki su arasında kılcal iletim boruları aracılığıyla devamlılık gösteren bir su sütununun oluştuğu düşünülürse, suyun yukarı doğru nasıl çekildiği kolayca anlaşılabilir.
Yapılan araştırmalar suyun gerilim direncinin, ksilem içindeki su sütununu en yüksek ağacın tepesine taşımaya yetecek kadar yüksek olduğunu göstermiştir. Bu mekanizma, boyları 100 m’yi geçen dev sekoya ağaçlarında bile suyun taşınmasını sağlayabilmektedir. Yani suyun yukarı doğru taşınmasında en fazla kabul edilen görüş terleme - çekim teorisidir.
Kılcallık: İnce bir hidrofilik (suya karşı çekim gösteren) tüp içinde, suyun yükselme eğilimi kılcallık olarak adlandırılır. Bu durumu basit bir deneyle göstermek mümkündür. Çapları farklı cam tüplerin uçları belirli bir hacimdeki suya batırılırsa, tüplerdeki su seviyeleri dıştaki sudan daha yüksek bir seviyeye ulaşır. İç yüzeyle ilişki halindeki su molekülleri yukarı doğru ilerlerken, hidrojen bağları ile bağlı oldukları diğer su moleküllerini de çekerler. Tüpün çapı genişledikçe, cam ile doğrudan temasta olan su molekülü yüzdesi düşer ve bu nedenle de suyun yükselişi az olur.
Bitkilerdeki ksilem de gözle görülemeyecek kadar ince kılcal borulardan meydana gelmiştir. Bu nedenle ksilem çeperlerinin su moleküllerini çekerek suyun yukarı doğru yükselmesini sağladığı düşünülmektedir. Ancak yapılan araştırmalar, suyun yükselmesinde kılcallığın diğer faktörlere göre daha etkisiz olduğunu göstermiştir.
Suyun Taşınmasında Stomaların Rolü
Bazı bitkiler (örneğin Elodea) tamamen su içinde yaşar. Bunlar suyu bütün yüzeyleriyle alırlar. Ancak çoğunluğu oluşturan kara bitkileri suyu topraktan kökleri yardımıyla almak zorundadırlar. Köklerle alınan su, iletim dokusuna ve yaprak damarlarına doğru hareket eder. Buradan da mezofildeki hücrelere geçerek buharlaşır. Buharlaşan bu su, en sonunda stomalardan dışarı atılır.
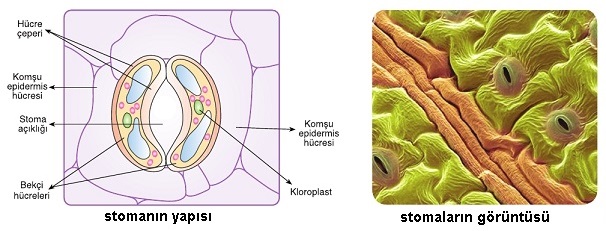
Epidermis hücrelerinin farklılaşmasıyla oluşan stomalar, bitkide terleme ve gaz alışverişi olaylarını düzenler. Bir stoma; bekçi ya da kilit hücresi de denilen iki adet stoma hücresi ile bunların aralarındaki stoma açıklığından (stoma porundan) oluşur. Stoma hücrelerinin önemli bir özelliği kloroplast içermeleridir. Bunların yanlarında bulunan epidermis hücreleri komşu hücreler adını alır. Komşu hücrelerde kloroplast bulunmaz.
Stomaların sıklık dereceleri, alt ve üst epidermisteki miktarları ve derinlikleri ile çevrenin ekolojik koşulları arasında bir ilişki vardır. Kurakçıl bitkilerde terlemeyle kaybedilecek su fazla değildir ve bu yüzden yaprak yüzeyindeki stoma sayısı azdır. Buna karşılık sucul ve nemcil bitkiler daha çok su alırlar ve bu bitkilerde suyun fazlasını atabilmek için daha çok stoma bulunması gerekir. Yaprakları su üstünde yüzen bitkilerin alt yüzeyi su ile temasta olduğundan, stomalar sadece üst epidermiste yer alır. Tamamen su altında kalan yapraklarda stoma bulunmaz. Kurakçıl bitkilerde stomalar iyice derine gömülmüşlerdir. Bu durum stomayı sıcak ve kuru rüzgârlardan koruyarak terleme hızını azaltır. Stoma çevresindeki tüyler de rüzgarı keserek stomanın yer aldığı çukurun nemli kalmasını sağlar ve böylece terleme hızının azalmasına neden olur. Stomalar, suyun bol olduğu bölgelerdeki bitkilerde çıkıntı yaparak yüzeyden daha yüksekte bulunurlar.
Bitkilerin buharlaşma yoluyla su kaybetmesine terleme adı verilir. Terlemenin yaklaşık %90’lık bölümü stoma yoluyla, geri kalanı ise kütikuladan yapılır. Bitkiler her gün birim ağırlık başına hayvanlara göre 10-20 kat daha fazla su kaybederler. Bunun nedenlerinden biri bitkilerin fotosentez için geniş bir yaprak yüzeyine sahip olmaları ve bunun da terleme yoluyla su kaybını artırmasıdır. Diğer neden ise yapraklar yoluyla atmosferden karbondioksit alınırken önemli oranda su kaybedilmesidir. Karbondioksit, yaprağın mezofil hücrelerine suda çözünerek girebilir. Bu nedenle mezofil hücrelerinin nemli olması gerekir. Hem mezofil hücrelerinin nemli olması hem de karbondioksit girişi için stomaların açık olması, hava ile temas eden su moleküllerinin buharlaşmasına yol açmaktadır. Terleme hızı; güneşli, sıcak, kurak ve rüzgarlı havalarda çok fazladır. Çünkü bu faktörler suyun buharlaşmasını artırır.
Karasal bitkilerde terleme, yapraklara su sağlamasının yanı sıra kökler aracılığıyla alınan inorganik maddelerin gövde ve yapraklara taşınmasına yardım eder. Ayrıca terleme, çevredeki havaya göre yaprağın sıcaklığını düşürebildiği için soğutma işlevi de görür. Böylece yaprak içindeki sıcaklık, metabolik işlevlerde görev yapan enzimlerin yapısını bozacak kadar yükselmemiş olur.
Terleme ile kaybedilen suyun çoğu stomalardan geçtiği için, terlemenin düzenlenmesinde stomaların açılıp kapanma özelliği büyük önem taşır.
Stomaların Açılıp Kapanma Mekanizması: Stomaların açılıp kapanması bekçi hücrelerindeki turgor basıncı değişimiyle kontrol edilir. Turgor basıncının artması stomanın açılmasına, azalması ise kapanmasına neden olur. Bekçi hücreleri çift çenekli bitkiler ile bazı tek çenekli bitkilerde böbrek biçimindedir. Bu hücrelerin stoma açıklığına bakan iç çeperleri, epidermis hücrelerine bakan dış çeperlerine göre daha kalındır. Bekçi hücrelerinde turgor basıncı artınca hücre çeperi gerilir. Bu gerilme hücrenin ince çeper tarafında daha fazla, kalın çeper tarafında daha az olur. Dışa bakan çeperler fazla gerildiği için, kalın çeperleri hücrenin içine doğru çekerek konkav (iç bükey) durum almasına yol açar. Böylece bekçi hücreleri arasındaki açıklık artar. Bekçi hücreleri su kaybettiğinde ise bu hücrelerin dışa doğru yaptığı kavis azalır ve stomalar kapanır.
Stomalar genel olarak gündüz açık geceleri kapalıdır. Gündüz bekçi hücrelerinde fotosentezle glikoz yoğunluğu artar ve komşu hücrelerden bekçi hücrelerine aktif olarak potasyum iyonları (K+) pompalanır. Çözünmüş maddedeki artış, bekçi hücrelerindeki su potansiyelini düşürür. Bunun sonucunda bekçi hücrelerine ozmos yoluyla su girer ve bu hücrelerin turgor basıncı artacağından stomalar açılır. Geceleyin olaylar tersine işler. Potasyum iyonları aktif olarak bekçi hücrelerinin dışına pompalanır. Bu durumda su ozmosla bekçi hücrelerinden dışarı çıkacağı için turgor basıncı düşer ve stomalar kapanır.
Çevresel faktörler, stomaların açılıp kapanmasını etkilemektedir. Normal olarak stomalar gündüz açık olmasına rağmen, su kıtlığı stomaların kapanmasına yol açabilir. Bitkiler su kıtlığı çekince mezofil hücrelerinde absisik asit olarak adlandırılan bir hormon üretilir. Bu hormon, bekçi hücrelerine stomaların kapatılması için uyarı gönderir. Stomaların kapanması bitkinin solmasını önlemesine rağmen, fotosentezin de yavaşlamasına neden olur.
Sıcak iklimlerdeki bazı bitkilerde gün ortasında bile kısa bir süre için stomalar kapanabilir. Bunun nedenlerinden biri, terleme yoluyla kaybedilen suyun, köklerle alınan sudan fazla olmasıdır. Bu durumda bekçi hücreleri turgorlu durumunu koruyamaz ve stomalar kapanır. Böylece bitki su miktarını korumuş olur. Stomaların kapanmasını sağlayan diğer bir neden ise mezofil dokusunda karbondioksit birikmesidir. Gün ortasındaki yüksek sıcaklıklar, solunum hızını artırır. Bu durum, yaprak içindeki karbondioksit konsantrasyonunu yükselterek stomaların kapanmasını uyarır. Stomaların kapanmasıyla bitki daha az su kaybeder ve solunum sonucu meydana gelen karbondioksit kullanılacağı için fotosentez durmaz.
Karanlık odada tutulan bir bitkide stomalar açılıp kapanarak günlük ritimlerini devam ettirirler. Bu durum stomaların açılıp kapanmasında bekçi hücrelerindeki içsel bir saatin de etkili olduğunu göstermektedir.
Fotosentez Ürünlerinin Taşınması
Bitkilerde fotosentez sonucu üretilen organik maddeler floem ile taşınır. Floemde en fazla taşınan şeker sakkaroz (sükroz) dur. Floem özsuyu ayrıca aminoasitler, hormonlar ve belirli iyonları da içerebilir. Taşıma işlemini özelleşmiş floem hücreleri olan kalburlu borular gerçekleştirir. Floemdeki taşıma ksilemdeki taşımaya göre daha yavaştır ve ksilemden farklı olarak çift yönlüdür. Fotosentez sonucu oluşan şeker, aminoasit gibi organik moleküller yukarıdan aşağıya doğru taşınırken, köklerde oluşan amino asit ve diğer azotlu organik moleküller de aşağıdan yukarıya doğru taşınmaktadır.
İletim demetindeki kalburlu borulardan biri floem özsuyunu bir yönde, aynı demetteki diğer bir kalburlu boru ise ters yönde taşıyabilir. Ancak bir tek kalburlu boruda aynı anda iki yönlü taşınım olmaz.
Organik maddelerin oluştuğu yer kaynak, kullanıldığı ya da depo edildiği yer ise havuz olarak adlandırılır. Kaynaklar genellikle olgun yapraklar, havuzlar ise kök ve meyve gibi organlardır. Bazı organlar farklı zamanlarda kaynak ya da havuz olabilir. Örneğin yabani şeker pancarının kökü ilk yılın büyüme mevsiminde yapraklardan (kaynaktan) gelen karbonhidratları biriktirdiği için bir havuz konumundadır. Aynı kök ikinci büyüme mevsiminde bir kaynağa dönüşür. Çünkü içerdiği nişasta şekere parçalanır ve şekerler de sürgün vermek için kullanılmak üzere gövdeye taşınır.
Fotosentez ürünlerinin (şekerlerin) kaynak konumundaki hücrelerden kalburlu boru elemanlarına geçişi “floemin yüklenmesi”, kalburlu boru elemanlarından havuz hücrelerine geçişi “floemin boşaltılması” olarak adlandırılır.
Organik maddelerin floemde taşınması “basınç akış teorisi” ile açıklanmaktadır.
Basınç - Akış Teorisi: Bu teori floemde taşınımın en çok kabul gören mekanizmasıdır. Bu olayın nasıl gerçekleştiği aşağıda özet halinde verilmiştir. (Aşağıdaki numaraları şekildeki numaralarla karşılaştırınız.)
Kaynaktan (bunlar çoğunlukla yaprak hücreleridir) ya da arkadaş hücrelerinden kalburlu borular içine şeker yüklenir. Bu durum kalburlu borularda derişimin artmasına yani su potansiyelinin azalmasına neden olur. Bunun sonucunda ksilemle kalburlu borular arasında derişim farkı ortaya çıkar ve su ozmosla ksilemden kalburlu borulara doğru girmeye başlar.
Kalburlu borulara giren su, bir sıvı basıncı (hidrostatik basınç) yaratır. Bu basıncın etkisiyle floem öz suyu kalburlu borular boyunca bir hücreden diğerine kütle halinde geçmeye zorlanır.
Kalburlu borulardaki şekerler derişim farkına bağlı olarak pasif ya da aktif taşımayla kök hücreleri gibi havuzlara boşaltılır. Böylece kalburlu borulardaki şeker derişimi azalır.
Şeker derişiminin azalması ile su kalburlu borulardan çıkarak ksileme geçer ve havuzdan kaynağa geri döner.
Sonuç olarak kalburlu borunun kaynak tarafında basıncın oluşması ve havuz tarafında basıncın düşmesi, kaynaktan havuza suyun ve bununla beraber şekerlerin taşınmasını sağlar. Suyun havuzdan kaynağa geri dönüşü trakeler aracılığıyla olur.