Hücresel Solunum: Enerjinin Açığa Çıkması
Canlı hücrelerde metabolik olayların çoğu için enerji gereklidir. Bu enerji, organik moleküllerin yıkımından elde edilir. Karmaşık yapılı organik moleküller önce enzimlerin faaliyeti ile daha küçük parçalara (yapı birimlerine) ayrılırlar. Örneğin; karbonhidratlar monosakkaritlere, yağlar yağ asitleri ve gliserole, proteinler ise aminoasitlere dönüştürülür. Bu olay insan ve hayvanların sindirim sisteminde gerçekleşir. Daha sonra kan yoluyla hücrelere taşınan bu yapı birimleri enzimlerin kontrolündeki bir dizi tepkime sonucu yıkılıp kimyasal bağ enerjileri açığa çıkarılır. Açığa çıkan bu enerjiden de ATP sentezlenir. Organik moleküllerin hücrede yıkılarak enerji (ATP) elde edilmesi olayına hücresel solunum denir.
Hücresel solunum olayı oksijenin kullanılıp kullanılmamasına göre oksijenli solunum ve oksijensiz solunum olarak ikiye ayrılır.
Oksijenli Solunum (Aerobik Solunum)
Oksijen, yerkabuğunun %50’sini teşkil eder. Ayrıca havada, suda, oksitlerde ve organik maddelerin yapısında bulunur. Atmosferde %21 oranında oksijen bulunmaktadır. Bu oksijenin kaynağını fotosentez sonucu ortaya çıkan serbest oksijen oluşturur. Denizlerdeki bitkisel planktonlar ve karalardaki bitkiler fotosentez yaparak atmosfere oksijen verir.
Oksijen, ozon tabakasının yapısında da bulunur. Ozon tabakası, atmosferden 20-50 km yükseklikte bulunan bir gaz katmanıdır. Bu katman, güneşten gelen ve canlıları olumsuz yönde etkileyen ultraviyole (morötesi) ışınları emerek yeryüzüne ulaşmasını engeller.
Günlük yaşamda solunum, soluk alıp verme anlamında kullanılır. Soluk alıp verme, oksijence zengin havanın akciğerlere girmesi, karbondioksitçe zengin havanın akciğerlerden dışarı çıkmasıdır. Oysa solunum kavramı hücresel bir olaydır. Oksijenli solunumda hücreye giren oksijen, besin maddelerini parçalar ve bu maddelerdeki kimyasal enerji hücrenin kullanabileceği ATP enerjisine dönüşür.
Hücredeki organik maddelerin oksijen varlığında karbondioksit ve suya kadar parçalanıp ATP elde edilmesine oksijenli solunum denir.
Oksijenli solunumu, hücrelerinde mitokondrisi olan bütün canlılar gerçekleştirir. Ayrıca prokaryot canlıların bazıları da oksijenli solunum yapar. Ancak oksijenli solunum yapan prokaryot canlılarda mitokondri bulunmadığından, bu olayı katalizleyen enzimler hücre zarının özel kıvrımlı bölgelerinde yer alır.
Hücre içinde organik moleküllerin parçalanması, her bir basamakta ayrı bir enzimin görev yaptığı tepkime zinciriyle gerçekleşir. Solunumda enerji kaynağı olarak karbonhidratlar, yağlar ve proteinler kullanılabilir. Canlılarda enerji kaynağı olarak öncelikle karbonhidratlar kullanıldığından, solunumda tepkimeye girecek organik bileşik olarak glikoz ele alınacaktır.
Oksijenli solunum glikoliz, krebs döngüsü ve elektron taşıma sistemi olarak üç aşamada gerçekleşir.
Glikoliz
Glikozun çeşitli enzimler kullanılarak pirüvik aside (piruvata) kadar yıkılması olayıdır. Glikoliz, prokaryot ve ökaryot hücrelerin sitoplâzmasında gerçekleşir.
Oksijen olsun ya da olmasın glikoliz reaksiyonları bütün canlılarda aynı şekilde gerçekleşir. Çünkü bu reaksiyonları etkileyen enzimler bütün canlılarda aynıdır.
Glikoliz, bazı ökaryot hücrelerin yeterli oksijen bulamadığı kritik durumlarda acil ATP gereksinimini karşılar. Ayrıca memelilerin alyuvar hücresi gibi bazı doku ve hücreler için tek metaboiik enerji kaynağıdır.
Glikoliz reaksiyonları şöyle özetlenebilir:
- Glikozun aktifleşmesi için önce bir molekül ATP’den bir fosfat grubu enzim aracılığıyla glikoza bağlanır ve glikoz-6- fosfat bileşiği oluşur (Oluşan bu bileşik, altıncı karbon atomuna bir fosfat grubu bağlanmış glikoz anlamına gelmektedir)
- Glikoz-6-fosfat, bir enzim yardımıyla fruktoz-6-fosfata dönüşür.
- Fruktoz-6- fosfat, bir molekül ATP’den bir fosfat grubu daha alınmasıyla fruktoz-1,6-difosfatı oluşturur. Buraya kadar olan reaksiyonlarda 2 ATP harcanmış ve şeker molekülü aktifleştirilmiştir.
- Fruktoz-1,6- difosfat bileşiği bir enzim yardımıyla her biri birer fosfat grubu taşıyan 3C’lu iki ayrı moleküle parçalanır. Oluşan yeni bileşikler fosfogliser aldehit (PGAL) olarak adlandırılır. PGAL, hem glikoliz hem de fotosentezde kilit rolü olan bir ara bileşiktir.
- Hidrojen ve elektron tutucu bir koenzim olan NAD+ (Nikotinamid Adenin Dinükleotit), PGAL’den iki hidrojen alınca NADH + H+ olarak indirgenir. Ayrıca her bir PGAL’e birer inorganik fosfat (Pi) grubu bağlanarak difosfogliserik asit (DPGA) oluşur. Bağlanan bu fosfatlar hücre sitoplâzmasındaki düşük enerjili fosfatlardan elde edilir. Ancak, bu düşük enerjili fosfatların PGAL’e bağlanırken enerjisi yükseltildiğinden, 3C'lu moleküllerin içerdiği yüksek enerjili fosfat grubu ikiye çıkmış olur. Bu yüksek enerjili fosfatlar ATP sentezinde kullanılacaktır.
- DPGA’ların birer fosfatı ADP’lere aktarılarak 2 ATP sentezlenir. Böylece iki molekül fosfogliserik asit (PGA) oluşur.
- 3C’lu birer fosfatlı PGA’ların, fosfatlarını ADP’lere aktarmasıyla 2 ATP daha sentezlenir. Böylece fosfat grubunu kaybeden PGA’lardan 2 molekül pirüvik asit (pirüvat) oluşur.
Glikolizde, substratların yapısındaki yüksek enerjili fosfatlar koparılarak doğrudan ADP’ye aktarılır ve ATP sentezi gerçekleşir. Buna substrat düzeyinde fosforilasyon denir.
Glikolizin başlangıcında bir glikoz molekülünün parçalanması için 2 molekül ATP harcanmış, ancak 4 molekül ATP sentezlenmiştir. Bu nedenle glikolizin net enerji kazancı 2 ATP’dir.
Glikoliz olayında bir molekül glikozdan; net 2 ATP, 2 NADH + H+ ve 2 pirüvik asit meydana gelir.
Krebs Döngüsü (Sitrik Asit Döngüsü)
Glikolizin son ürünü olan 3C’lu pirüvik asit, ortamda oksijen bulunduğu zaman sitoplâzmadan mitokondriye girer. Pirüvik asitten sonraki parçalanma olayları hücrenin mitokondrisinde gerçekleşir.
Mitokondriye giren pirüvik asitten bir molekül karbondioksit ve iki hidrojen atomu ayrılır. Ayrılan hidrojenler NAD+ koenzimi tarafından tutulur ve NADH + H+ oluşur. Bunun sonucunda pirüvik asit, krebs döngüsünü başlatacak temel madde olan Asetil-CoA’ya dönüşür.
Pirüvik asit mitokondride Asetil-CoA’ya dönüşürken bir CO2 açığa çıkmakta ve NADH + H+molekülü oluşmaktadır. Ancak glikoliz reaksiyonları sonucu iki molekül pirüvik asidin meydana geldiği unutulmamalıdır. Bu nedenle yukarıdaki reaksyonda açığa çıkan C02 ve oluşan NADH + H+ sayısı iki olarak düşünülmelidir.
Krebs döngüsü reaksiyonları ilk kez Hans KREBS tarafından bulunduğu için krebs döngüsü adını alır. Bu reaksiyonlar mitokondrinin matriksinde gerçekleşir. Krebs döngüsü aşağıda bazı ara basamaklar verilmeden özetlenmiştir.
- Krebs döngüsünde ilk olarak iki karbonlu Asetil-CoA, dört karbonlu bir molekül olan okzaloasetik asitle blrleşerek altı karbonlu sitrik asidi oluşturur.
- Sitrik asit, beş karbonlu bir bileşiğe dönüşürken bir molekül karbondioksit açığa çıkar. Ayrıca bu basamakta ayrılan iki hidrojen atomu, NAD+ tarafından tutulur ve NADH + H+ oluşur.
- Beş karbonlu bileşikten de bir molekül karbondioksit ile iki hidrojen atomu ayrılır ve hidrojenler NAD+ tarafından tutularak NADH + H+ meydana gelir. Sonuçta dört karbonlu bir bileşik oluşur.
- Dört karbonlu bileşik başka bir dört karbonlu bileşiğe dönüşürken substrat düzeyinde fosforilasyonla bir ATP üretilir.
- Daha sonra bu bileşikten iki hidrojen atomu ayrılır ve FAD (Flavin Adenin Dinükleotit) koenzimi tarafından tutularak FADH2 oluşur. Böylece yeni bir dört karbonlu bileşik meydana gelir.
- Son basamakta dört karbonlu bileşik iki hidrojen atomu daha kaybeder. Bu hidrojenler NAD+ tarafından tutularak NADH + H+ oluşur. Böylece başlangıçta Asetil-CoA ile reaksiyona giren dört karbonlu okzaloasetik asit yeniden meydana gelir.
Dört karbonlu bileşik yeni bir Asetil-CoA ile reaksiyona girer ve krebs döngüsü sürdürülür.
Anlatılan bu reaksiyonlarda bir molekül Asetil-CoA’nın krebs döngüsüne katılması gösterilmiştir. Ancak daha önce açıklandığı gibi krebs döngüsü iki koldan yürütülmektedir ve bu nedenle döngüye iki molekül Asetil-CoA girmektedir.
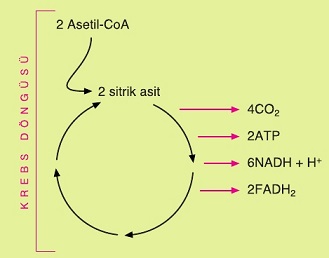
Krebs döngüsüne iki molekül Asetil-CoA girdiği zaman; 4 CO2 açığa çıkmakta, substrat düzeyinde fosforilasyonla 2 ATP sentezlenmekte, 6NADH + H+ ve 2 FADH2 oluşmaktadır.
Elektron Taşıma Sistemi (ETS)
Elektron taşıma sisteminin daha iyi anlaşılabilmesi için yükseltgenme (oksidasyon) ve indirgenme (redüksiyon) kavramlarının bilinmesi gereklidir.
Oksijenli solunumda görev yapan ETS elemanları ökaryot hücrelerde mitokondrinin iç zarında, prokaryot hücrelerde ise hücre zarında bulunur. Bu safhada ATP sentezi yükseltgenme ve indirgenme reaksiyonlarıyla sağlanır.
NAD+ve FAD, solunumda görev yapan koenzimlerdir. Bu koenzimlerin yapılarında vitamin bulunur. Örneğin, hücreler NAD+'ı B3 vitamininin bir türevi olan nikotinamidden sentezlerler. FAD koenzimi ise B2 vitamini içerir. NAD+'ın indirgenmiş hali NADH, FAD'ın indirgenmiş hali ise FADH’dır.
NAD koenzimine, bir çift hidrojen atomu aktarıldığında, 2 elektronla bir proton tutar ve bir proton serbest hale geçer. Bu durum NADH + H+ halinde gösterilir. FAD koenzimi ise iki hidrojen atomunu tam olarak tutar. Bu durum da FADH2 halinde gösterilir.
Elektron taşıma sistemi oksijenli solunumda enerjinin en yoğun üretildiği safhadır. Mitokondrinin iç zarı kristaları oluşturacak şekilde kıvrımlar yapmıştır. Bu zar elektron taşıma sistemi ve ATP sentaz enzimini taşır. ETS’deki ATP sentezi kemiozmotik hipotezin ortaya konmasından sonra açıklanmıştır.
Bu hipoteze göre ETS'deki olayları özetleyelim:
Glikoliz ve krebs döngüsünde oluşan NADH + H+ ve FADH2’deki hidrojen atomlarının protonları ve yüksek enerjili elektronları, elektron taşıma sistemine geçerler. Koenzimlerle taşınan elektronlar taşıyıcı moleküllere aktarılırken protonlar matrikse bırakılır. Bu sırada NADH + H+ ve FADH2, sırasıyla NAD+ ve FAD formlarına yükseltgenir. Böylece NAD+ ve FAD molekülleri tekrar elektronları kabul edebilecek hale gelir.
Mitokondrinin iç zarında yer alan ETS elemanları NADH-Q redüktaz, ubikinon, sitokrom redüktaz, sitokrom c ve sitokrom oksidazdır (Ubikinon dışındaki ETS elemanları protein yapısındadır).
Taşıma sistemindeki elektronların enerji seviyeleri, kademeli olarak düşürülür. Bu sırada açığa çıkan serbest enerjinin bir kısmı ısı olarak ortama yayılır. Enerjinin büyük bir kısmı ise, protonların matriksten mitokondrinin iç ve dış zarı arasında bulunan boşluğa pompalanmasında kullanılır.
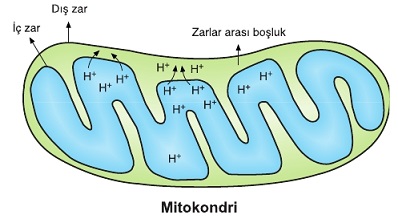
Böylece mitokondrinin iç ve dış zarı arasındaki boşluk matrikse oranla daha yüksek proton derişimine sahip olur. Protonlar elektrik yükü taşıdığından iç zarın iki yüzü arasında elektriksel yük farkına da yol açar. Bu derişim ve elektrik yük farkı bir potansiyel enerji oluşturur.
Mitokondrinin iç zarı protonlara geçirgen değildir. ATP sentaz enzimi zarlar arası boşluktaki protonların tekrar matrikse akışını sağlayan bir yol oluşturur. ATP sentaz enzimi proton akışı sırasında aktifleşir ve enzimde yapısal değişimler oluşur. Bu değişimler ADP’ye bir fosfat eklenerek ATP sentezlenmesini sağlar. Protonlar matrikse doğru aktıkça ATP sentezi de devam eder.
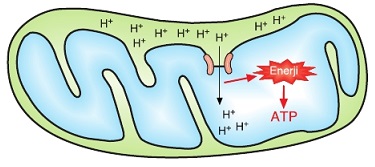
ETS'nin son elektron alıcısı oksijendir. Elektron taşınmasının son aşamasında düşük enerjili hale gelmiş olan elektronlar, oksijen ve protonlarla (H+) birleşerek suyu oluşturur. Oksijenli solunumda ETS aracılığıyla ATP sentezlenmesine oksidatif fosforilasyon denir.
Protonlar ve elektronlar NAD tarafından (NADH + H* şeklinde) ETS’ye iletilirse 3 ATP sentez- lenir. Eğer protonlar ve elektronlar FAD tarafından alınıp (FADH2 şeklinde) ETS’ye taşınırsa 2 ATP sentezlenir.
Solunumda enerjinin büyük bölümü glikoliz ve krebs döngüsünde açığa çıkan hidrojen atomuna ait elektronların ETS’de taşınması ile elde edilir.
Buraya kadar anlatılan oksijenli solunum reaksiyonlarını şöyle özetleyebiliriz:
Reaksiyonlar sırasında glikozdan basamak basamak karbon ve hidrojen atomları koparılır. Karbon atomları, karbondioksit olarak dışarı verilir (Karbondioksitteki oksijenlerin glikozdan geldiğine dikkat edilmelidir). Hidrojen atomlarının yüksek enerjili elektronları ise mitokondri iç zarındaki elektron taşıma sisteminde (ETS), oksijene doğru taşınır. Elektronlar bu taşınım sırasında giderek daha düşük enerji düzeyine geçerler. Elektron taşıma zinciri yoluyla elde edilen enerji, sonradan ATP üretiminde kullanılır. Buna oksidatif fosforilasyon adı verilir. Elektronların enerji düzeyi, oksijene ulaştığında en düşük seviyededir. Elektron alan oksijen, hidrojenin protonuyla birleşerek suyu oluşturur. (Sudaki oksijenin, glikozdan değil oksijen molekülünden geldiğine dikkat edilmelidir.)
C6H12O6 + 6O2 → 6CO2 + 6H2O + 38 ATP
Sonuç olarak oksijenli solunumda CO2 ile H2O oluşur ve ATP üretilir. Şimdi bu moleküllerin oluşumunu ayrı ayrı inceleyelim.
ATP üretimi: Oksijenli solunumda hem substrat düzeyinde fosforilasyonla hem de oksidatif fosforilasyonla ATP üretilir.
Substrat düzeyinde fosforilasyonla ATP üretimi
- Glikoliz safhasında 4 ATP sentezlenir. 2 ATP aktivasyon enerjisinde harcandığından ATP kazancı 2’dir.
- Krebs çemberinde 2 ATP sentezlenir.
Oksidatif fosforilasyonla ATP üretimi
- Glikoliz safhasında 2 NADH + H+ molekülü oluşur. 2H atomu NAD tarafından ETS’ye girdiğinde 3 ATP sentezlendlğine göre, 2 NADH + H+ daki 4H atomu 6 ATP sentezine olanak sağlar.
- Pirüvik asidin Asetil-CoA’ya dönüşmesi sırasında oluşan 2 NADH + H+’daki 4H atomu kullanılarak ETS’de 6 ATP sentezlenir.
- Krebs döngüsünde 6 NADH + H+ oluşur. 6 NADH + H+’daki 12H atomu 18 ATP sentezine olanak sağlar. Krebs döngüsünde ayrıca 2 FADH2 meydana gelir. 2H atomu FAD tarafından ETS’ye girdiğinde 2 ATP sentezlendiğine göre, 2 FADH2’deki 4H atomu 4 ATP sentezine yol açar.
Bir molekül glikozun CO2 ve H2O’ya parçalanmasından toplam 40 ATP, net 38 ATP sentezlenir. 40 ATP’nin 6 tanesi substrat düzeyinde fosforilasyonla, 34 tanesi ise oksidatif fosforilasyonla üretilir. Substrat düzeyinde elde edilen 6 ATP’nin 4 tanesi sitoplâzmada (glikolizde), 2 tanesi mitokondride (krebs döngüsünde) sentezlenir. Oksidatif fosforilasyonla elde edilen 34 ATP’nin tamamı mitokondride sentezlenir.
Suyun oluşumu: Oksijenli solunumda, ETS’ye aktarılan bir çift hidrojen atomunun oksijen ile birleşmesi sonucu 1 molekül H2O oluşur. NAD ve FAD’ın tuttuğu hidrojen atomu sayısı 24’tür. Bu 24 hidrojen atomunun oksijen ile birleşmesi sonucu 12 molekül H2O meydana gelir.
CO2 oluşumu: Glikoliz sonucu oluşan pirüvik asidin, krebs döngüsünü başlatacak temel madde olan Asetil-CoA’ya dönüşümü sırasında bir molekül CO2 açığa çıkar. Krebs döngüsünde ise 6C’lu sitrik asitten 5C'lu bileşiğin oluşumunda ve 5C’lu bileşikten 4C’lu bileşiğin oluşumunda birer molekül CO2 çıkışı olur.
Buna göre 1 molekül pirüvik asidin tepkimeye girmesi sonucu 3 molekül CO2 açığa çıkar. Glikoliz sonucu 2 molekül pirüvik asit oluştuğu için toplam 6 molekül CO2 çıkışı olur. Oksijenli solunumda karbondioksitlerin tamamının mitokondride açığa çıktığını bilmek gerekir.
Solunumda Enerji Verimi
Oksijenli solunum bir yanma olayı değildir. Eğer bir molekül glikoz kalorimetrede oksijenli olarak yakılırsa CO2 ve H2O açığa çıkar ve enerji ısı olarak çevreye yayılır. Ancak glikozun hücrede oksijenli solunumla yıkımı sırasında enerjinin yaklaşık %40’ı ATP'de tutulur (Yani oksijenli solunumun enerji verimi yaklaşık %40'tır). Geriye kalan enerjinin bir kısmı ısı enerjisine dönüşür. Isı enerjisi kullanılamayan bir enerji gibi düşünülse de vücut sıcaklığının düzenlenmesinde rol oynar. Enerjinin bir kısmı da oksijenli solunumun son ürünleri olan H2O ve CO2 moleküllerinin bağları arasında kalır.
Oksijensiz solunum yapan bir hücrenin enerji verimi %2 - 10 arasındadır.
Organik Besinlerin Oksijenli Solunumda İzlediği Yol
Hücreler ATP enerjisini sadece karbonhidratlardan değil yağlardan ve proteinlerden de sağlarlar. Besin maddelerinin oksijenli solunumla metabolik olarak parçalanma reaksiyonları çok farklı değildir. Krebs döngüsü besinlerin ortak yıkım yoludur.
Yağlar önce hidrolize uğrayarak gliserol ve yağ asitlerine dönüşür. Daha sonra üç karbonlu bir bileşik olan gliserol, PGAL’e dönüştürülerek glikolitik yola sokulur. PGAL'den de pirüvik asit oluşur. Yağ asitleri ise Asetil-CoA’ya dönüştürülerek solunum tepkimelerine katılır.
Yağlar önce hidrolize uğrayarak gliserol ve yağ asitlerine dönüşür. Daha sonra üç karbonlu bir bileşik olan gliserol, PGAL’e dönüştürülerek glikolitik yola sokulur. PGAL'den de pirüvik asit oluşur. Yağ asitleri ise Asetil-CoA’ya dönüştürülerek solunum tepkimelerine katılır.
Yağlar karbonhidratlara göre daha yüksek oranda hidrojen içerdiklerinden, tam olarak oksitlendiklerinde birim ağırlık başına daha fazla enerji oluştururlar. Bir gram yağ, bir gram karbonhidratın verdiği enerjiden iki kat daha fazla enerji verir.
Proteinlerin hidrolizi sonucu oluşan aminoasitler bir dizi yolla parçalanırlar. Yıkım sırasında önce amino grubu, amonyak (NH3) halinde ayrılır. Oluşan amonyaktan karaciğerde üre sentezlenir. Amino grubunu kaybeden aminoasitler karbon sayılarına göre pirüvik aside, Asetil-CoA’ya veya krebs döngüsü bileşiklerine dönüştürülür.
Bir gram proteinin tam parçalanması sonucu yaklaşık olarak bir gram karbonhidratın verdiği enerjiye eşit miktarda enerji açığa çıkar.
Karbonhidrat ve yağlar solunum tepkimelerinde kullanıldığında son ürün olarak C02 ve H20 açığa çıkarken proteinlerin kullanıldığı durumlarda CO2, H2O ve NH3 oluşur.
Oksijensiz Solunum (Anaerobik Solunum = Fermantasyon)
Organik maddelerin oksijensiz ortamda parçalanarak enerji elde edilmesi olayına fermantasyon denir. Oksijen yokluğunda ya da yetersizliğinde oksidatif fosforilasyon gerçekleşemez. Bu koşullardaki hücrede sadece glikoliz evresinde substrat düzeyinde fosforilasyonla ATP sentezlenir. Çünkü glikoliz evresinde, glikozu pirüvik aside parçalayan kimyasal reaksiyonlar oksijen gerektirmez.
Oksijenli solunumda glikoz başına net 38 ATP üretilmesine karşılık, fermantasyonda bu rakam net 2 ATP'dir. Enerji veriminin düşük olmasına rağmen, fermantasyon birçok canlıya oksijen eksikliğinde yaşayabilme olanağını sağlamak-
1857 yılında Fransız kimyageri L. Pasteur fermantasyonun canlı maya hücreleri tarafından gerçekleştirildiğini bulmuştur. 1907 Nobel Kimya Ödülü'nü kazanan E. Buchner fermantasyonun canlı hücrelere özgü bir olay olmadığını, maya hücrelerinin parçalanması sonucu elde edilen özsuyun da fermantasyon gücüne sahip olduğunu göstermiştir. Daha sonraki yıllarda yapılan çalışmalar, bu sıvıda fermantasyonun gerçekleşmesini sağlayan çeşitli enzimlerin bulunduğunu göstermiştir (Enzimlerin hücre dışında da görev yaptığını hatırlayınız).
İnsanlar, bazı canlılar tarafından gerçekleştirilen fermantasyon olaylarından çeşitli şekillerde faydalanmaktadır. Örneğin maya hücreleri ve diğer bazı mikroorganizmalar tarafından yapılan fermantasyon sonucu ekmek ve alkollü içecekler elde edilmektedir. Yine bakterilerin yaptığı fermantasyonlar; peynirlerin büyük bir kısmının, yoğurdun ve bazı ürünlerin üretiminde önem taşır.
Fermantasyon reaksiyonlarında görev yapan enzimler hücrenin sitoplâzmasında bulunur.
Daha önce de açıklandığı gibi glikoliz reaksiyonları oksijenli ve oksijensiz solunumda ortak gerçekleşir. Glikoliz evresinde bir glikoz molekülü sitoplâzmada 2 pirüvik aside parçalanır. Başlangıç reaksiyonlarında 2 ATP harcanmasına karşılık, sonraki reaksiyonlarda 4 ATP üretildiği için 2 ATP kazanılmış olur. Ayrıca 2 NAD+ molekülü 2 NADH + H+ şeklinde indirgenir.
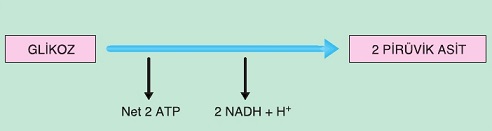
Ortamda oksijen olmadığı için Asetil-CoA’ya dönüşemeyen pirüvik asit; etil alkol, laktik asit, bütanol, aseton, bütirik asit gibi ürünlere dönüşebilir. Son ürün farklılığı enzim farklılığına dayanır.
Fermantasyon reaksiyonları sonucu oluşan maddeler, oksijen yokluğunda daha fazla parçalanamadığından kimyasal enerji içerirler ve bunlar hücre için artık üründürler. Bu yüzden fermantasyonla elde edilen A TP miktarı, oksijenli solunuma göre daha azdır.
Fermantasyon çeşitleri ürettikleri ürünlere göre adlandırılır. Bunların en önemlileri etil alkol ve laktik asit fermantasyonlarıdır.
Etil Alkol Fermantasyonu
Glikozdan etil alkolün oluştuğu ve enerjinin açığa çıktığı reaksiyonlar dizisine etil alkol fermantasyonu denir.
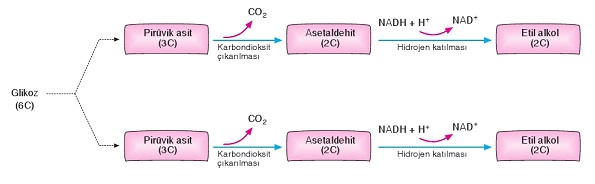
Etil alkol fermantasyonunda glikoliz sonucu oluşan pirüvik asitlerin her birinden birer karbondioksit çıkarak 2 molekül asetaldehit meydana gelir. Asetaldehitlere de glikoliz evresinde oluşan NADH + H+’lardaki hidrojenlerin katılmasıyla 2 molekül etil alkol (C2HgOH) oluşur. Böylece NAD+ koenzimleri yeniden kullanılmak üzere serbest kalır.
Etil alkol fermantasyonu, başta bira mayası olmak üzere maya mantarlarında, bazı tohumlarda ve şarap bakterilerinde gerçekleşen bir enerji elde etme yöntemidir. Üzüm suyundan şarap oluşumu alkolik fermantasyona örnek olarak verilebilir. Fermantasyon reaksiyonları sonucu açığa çıkan alkol oranı %18’i geçerse hücreler üzerinde zehir etkisi yapar.
Laktik Asit Fermantasyonu
Glikozdan laktik asidin oluştuğu ve enerjinin açığa çıktığı reaksiyonlar dizisine laktik asit fermantasyonu denir.
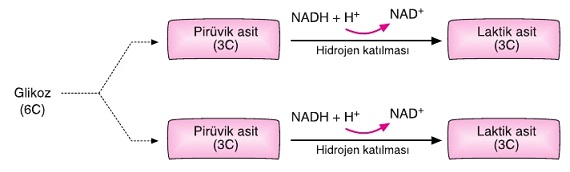
Laktik asit fermantasyonunda glikoliz sonucu oluşan pirüvik asitlerin her birine NADH + H+'lardaki hidrojenler katılır. Böylece 2 molekül laktik asit (C3H6O3) oluşur.
Aşağıda laktik asit fermantasyonunun denklemi gösterilmiştir.
C6H12O6 + 2ATP → 2C3H6O3 + 4ATP
Laktik asit fermantasyonunda, etil alkol fermantasyonundan farklı olarak karbondioksit açığa çıkmaz.
Laktik asit, glikolizin son ürünlerinin birikimini önleyen bir çeşit sigortadır. Laktik asidin oluşumu, pirüvik asit ve NADH + H+,lardaki hidrojenlerin ortamdan uzaklaştırılmasını sağlar. Eğer bu dönüşüm olmasaydı, glikoliz ancak birkaç saniye daha devam edebilir ve ATP üretimi dururdu.
Pirüvik asidin etil alkol veya laktik aside (fermantasyon ürünlerine) dönüşmesi ATP üretilmesine yol açmaz. Ancak bu süreç fermantasyon için çok önemlidir. Çünkü glikoliz sırasında indirgenerek NADH haline geçen NAD’ın tekrar yükseltgenmesini ve böylece glikolizin devam etmesini sağlar.
Oksijenli solunumda NAD+ ya da FAD’a aktarılan hidrojenler, elektron taşıma sisteminde oksijene kadar taşınır ve su oluşur. Ancak oksijensiz solunumda NAD*’a aktarılan hidrojenler fermantasyon ürünlerinin oluşumunu sağladığı için su meydana gelmez.
Laktik asit fermantasyonu bakteri ve hayvan hücrelerinin bazılarında görülen bir enerji elde etme yöntemidir.
İnsan Vücudunda Laktik Asit Fermantasyonu
Çizgili kas hücreleri yeterli oksijen elde edemediği zaman laktik asit fermantasyonuyla enerji üretmeye devam ederler. Az miktarda laktik asit, kasın daha iyi çalışmasını sağlar. Spora başlamadan önce ısınma hareketleri yapılmasının nedeni budur. Çizgili kasların fazla çalışması sonucu meydana gelen laktik asit, kan dolaşımı yoluyla beyne ulaşarak yorgunluk ve uyku duygusunu meydana getirir. Yoğurtta da bol miktarda laktik asit bulunur. Yoğurt yenildiğinde uykunun gelme sebebi budur.
Kasta oluşan laktik asidin bir kısmı burada kalırken çoğu kan yoluyla karaciğere taşınır. Aşırı aktivite sona erdikten sonra hızlı soluk alıp vermelerle karaciğer ve kas hücrelerine yeterli oksijen ulaşır. Bunun sonucunda laktik asit oluşturan tepkimeler tersine döner ve laktik asit, pirüvik aside dönüşür. Pirüvik asit de oksijenli solunuma katılır. Karaciğer hücreleri laktik asidin bir kısmını pirüvik aside dönüştürürken, geri kalanını glikoza çevirir ve bundan da glikojen sentezlenir.
Kas kasılması için gerekli enerji ATP'den sağlanır. Yoğun aktivite sırasında ATP, gerekli enerjiyi ancak 0,5 saniye kadar karşılayabilir. Kasılma sırasında harcanan ATP’nin en kısa yoldan sentezi için kreatin fosfat molekülü kullanılır.
Dinlenme sırasında kreatin molekülü ATP’den bir fosfat alarak kreatin fosfat haline gelir.
Kreatin + ATP → ADP + Kreatin fosfat
Kasılma sırasında ATP enerjisi kullanılıp bitince, kreatin fosfat hidroliz olarak fosfatını ADP'ye aktarır. Böylece ATP yeniden ve hızlı bir şekilde elde edilmiş olur.
Kreatin fosfat + ADP → ATP + Kreatin
Kreatin fosfat molekülü sadece çizgili kas hücrelerinde bulunur ve miktarı ATP’den 20 kat daha fazladır. Bu molekül kas kasılması için doğrudan enerji sağlamaz, fakat yüksek enerjili fosfat grubunu ADP’ye vererek ATP oluşmasına yol açar. ATP, kasılma için doğrudan enerji sağlayan moleküldür. Çizgili kas hücrelerinde harcanan ATP’nin yeniden üretilebilmesi için kullanılan moleküller sırasıyla; kreatin fosfat, glikoz ve glikojendir.
Kas kasılması sırasında; glikojen, glikoz, oksijen, ATP ve kreatin fosfatın miktarı azalır. Buna karşılık karbondioksit, kreatin, ADP, fosfat, laktik asit ve ısı miktarında artış olur.